ENTRAINMENTENTRAINMENT
ENTRAINMENTENTRAINMENT![]()
The daily rhythms of most plants and animals are synchronised to a period of exactly 24 hours, a process called entrainment. The clock synchronises using environmental time cues called Zeitgebers (from the German for "timegiver"). These are such diverse things as light, temperature and social activity.
Light
Light is the most obvious and well studies zeitgeber. In the natural world the light-dark cycle caused by the rotation of the Earth (i.e. day and night) provides a precise daily signal. At most latitudes the light-dark signal fluctuates seasonally in a precise way. In a laboratory, most organisms will readily entrain to an artificial 24 hour light-dark cycle.
Light is detected by photoreceptors. In some cases the receptor is obvious e.g. the retina in animals, and prevention of the detection of light by these receptors causes freerunning. However some naturally blind animals amongst the vertebrates can synchronise to light-dark cycles without the presence of obvious light detecting apparatus. This means that there may be other photoreceptors may be involved (Lucas et al 1999).
Temperature
Temperature changes may provide potential Zeitgebers. These temperatures occur in the natural world through the same mechanisms as light-dark cycles (i.e. the sun). However due to daily and seasonal changes, temperature cues are less precise. Never the less some organisms e.g. lizards can entrain to temperature cycles with a change of less than 1°C per cycle (Binkley 1997).
Social Cues
Social cues from other organisms can act as Zeitgebers. For example human volunteers kept in constant dark with 24 hour feeding and sleep cycles stayed entrained to 24 hours. (Binkley 1997). Submariners are entrained to an 18 hour day as most submarines operate on a 3 x 6-hour watch system. (Binkley 1997). The secretion of Luteinising hormone in male hamsters increases in the presence of female hamsters. However this only occurs after a critical photoperiod length (Anand et al 2002).
Limits of entrainment
A circadian rhythm can entrain to a 24 hour cycle quite easily. The length of the cycle providing the time cues is designated T. It is possible to entrain organisms to cycles with a T different to 24 hours.
As a rule there is a range in which an organisms can be entrained. It is possible to entrain an organism to 20 or 22 hours (LD 10:10 T=20, or LD 11:11 T=22). Below T=16, organisms will free run. The limits are different for different organisms. In mice and hamsters the limits are T=21 to T=26. In most other animals and plant is possible to entrain the organism down to T=18 hours and as much as 28 to 30 hours. (Bünning 1973).
Entrainment to a non 24 hour system can have a dramatic effect on behaviour. When night-active mice are entrained to a 25 hour cycle, their amount of activity and maximal activity levels are advanced compared to a 24 hour cycle, so that they almost become light-active. On the other hand, dark-active mice entrained to a 22 hour cycle are even more dark active (Aschoff 1965).
Sometimes organisms kept in exotic short cycles (e.g. LD 6.6, LD 3:3 or LD 2:2) do not entrain to these new cycles or freerun but instead remain entrained to a 24 hour cycle. This occurs when the external cycle has a period that is a multiple of the internal rhythm. For example, this might occur when the LD cycle has a frequency of 2 cycles/ 24 hours, or 3 cycles/ 24 hours (in other words had a period of 12 or 8 hours respectively. This phenomenon is known as frequency demultiplication.
Photoperiod
In a light-dark cycle, the period of light is called the photoperiod, and the period of dark is called the scotoperiod. The length of the photoperiod can affect entrainment. Sparrows can be entrained to extremes of 1 hour of light per 24 hour cycle or 23 hours of light per 24 hour cycle (Binkley 1997). However this affects behaviour. When there is less than 6 hours of light, the sparrows are active during the photoperiod only. However when there are less than 6 hours of light, the sparrows are active in the dark in anticipation of the light (Binkley 1997).
Phase angles
There can be a difference between the entraining cycle and the entrained rhythm. This difference can be measured by selecting a point in the entraining cycle (e.g. light on), and a phase reference point in the resulting rhythm (e.g. the onset of a particular activity). The difference between these two points is known as the phase angle. The phase angle is a function of photoperiod, for example it may be greater at LD 1:23 than at LD 4:20 (i.e. there may be a greater time between say light on and the start of activity at a shorter photoperiod).
In humans the phase angle difference can be seen between the natural lights on i.e. dawn, and waking up (particularly in students!).
Reentrainment
When we travel east, the sun rises and sets earlier. The natural light is advanced with respect to that at home. This results in an apparently shorter night followed by a new cycle. When we travel west, sunrise and sunset is later than at home, giving an apparently longer day, followed by a new cycle. In both these cases, we undergo a phase shift of the Zeitgebers in our environment.
Although north-south travel does not result in the crossing of time zones, there may still be photoperiod changes due to a change in latitude.
In the cases of both longitudinal and latitudinal travel, the internal circadian rhythms must reset so that they are synchronised with local environmental Zeitgebers. During the period of adaptation, intermediate cycles may be seen. These are called transient. The entrainments before and after phase shifts are called the steady state.
In humans, the period of readaptation during travel is called jet lag. However phase shifting does not exclusively occur with travel. The increased need for 24 hour service, or the constant use of expensive machines has resulted in the use of shift work to provide a constant work force. Such schedules often require the frequent resynchronisation of individuals to new time cues as a result of working shifts. For more detailed information, refer to the chapters on Resetting and Changes to the rhythm)
Phase shift notation
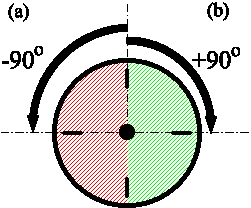
In the laboratory, changing the timing of entraining signals can stimulate phase shifts. The notation for denoting phase shifts is based on a clock with 24 hours = 360 degrees. If the phase shift delays the cycle it is given a minus sign; if it advances the cycle it is given a plus sign. The time by which the cycle is shifted is translated into degrees.
Both phase shifts correspond to a 6 hour shift. As 24 hours=360 degrees, 6 hours corresponds to 90 degrees. (a) The phase shift delays the cycle by 6 hours. (b) The phase shift advances the cycle by 6 hours.
A 180 degree shift (corresponding to a 12 hour phase shift) that completely reverses the times of light and dark in a 24 hour cycle is called a phase reversal.
The time taken to resynchronise to a phase reversal varies according to the organism. Cockroaches and sparrows readjust in 4 days whilst rats take 8 days. Humans take a full 11 days to completely resynchronise to a 12 hour phase shift. These times reflect not only the organism, but also the rhythm measures and the nature of the zeitgeber. Resynchronisation will take less time with a shorter phase shift (Aschoff et al 1975).
Incomplete readjustment
A zeitgeber needs to be at a particular strength in order to have an entraining effect. If the signal if at the threshold of the entrainment i.e. strong enough to catch a rhythm, but not strong enough to hold it, the result may be incomplete entrainment.
In relative coordination, the organisms’ rhythm is roughly synchronised by the cycle, but it drifts away from the signal then periodically regains it.
Sometimes and organism freeruns past a zeitgeber with a long period then freeruns with a short period to catch the signal. This is called bounce (Pittendrigh and Daan 1976).
Multiple signals
In the natural environment, there is often more than one zeitgeber acting at the same time. Whist some of these may reinforce each other e.g. light and temperature generated by the sun, the Zeitgebers may also give competing signals to the same rhythm. These are called conflicting Zeitgebers (Binkley1997)
In laboratory experiments fruit flies may be exposed to simultaneous light and temperature cycles with varying phase relationships between light and temperature. Fly eclosion (the emergence of an adult fly from a pupal case) followed the low part of the temperature cycle during the light time. However the rhythm could not follow the temperature cycle in the dark there was a 180 degree area in which the temperature cycle could not drive the rhythm. This is called the zone of forbidden phase (Binkley 1997). Flies however have body temperature that fluctuate with environmental temperature and hence are more dependent on the environment. Animals that maintain a constant body temperature are less affected by temperature cues.